ABSTRAKT
Vodní organismy mají velmi dobrou schopnost reflektovat podmínky prostředí, ve kterém žijí, proto jsou často využívány k posouzení ekologického stavu. Společenstvo juvenilních ryb (0+) představuje vhodný nástroj pro monitoring ekologického stavu vodních toků, protože vykazuje velmi rychlou reakci na měnící se podmínky. Cílem studie bylo posouzení stavu společenstva juvenilních ryb (0+) na 22 sledovaných lokalitách napříč Českou republikou (ČR) v letech 2019–2021. Odlov ryb byl prováděn elektrolovem (bateriovým agregátem) v mělkých úsecích podél břehové linie. Společenstvo juvenilních ryb (0+) bylo relativně bohaté, celkem bylo zaznamenáno 36 druhů, minimálně byly zachyceny čtyři a maximálně 15 druhů na lokalitu (průměrně se vyskytovalo devět druhů na lokalitu). Ve sledovaném období byly zjištěny významné rozdíly ve společenstvu juvenilních ryb (0+) napříč lokalitami i mezi jednotlivými lety. Zhodnocení ekologického stavu bylo realizováno pomocí českého multimetrického indexu (CZI). Byly zaznamenány zásadní změny ekologického stavu: čtyři lokality vykazovaly výrazné zhoršení a čtyři naopak odrážely výrazné zlepšení stavu. Na zbylých lokalitách byl stav spíše setrvalý (nedocházelo k výraznému zlepšení ani zhoršení). Nejlepší ekologický stav, kde společenstvo juvenilních ryb odpovídá přirozenému vodnímu toku, byl zjištěn na lokalitách Orlice v Nepasicích (Královéhradecký kraj) a Olše ve Věřňovicích (Moravskoslezský kraj). Naopak nejnižší hodnoty CZI vykazovaly lokality Ohře – Želina (Ústecký kraj), Dyje – Podhradí nad Dyjí, Dyje – Jevišovka (Jihomoravský kraj) a Cidlina – Sány (Středočeský kraj), které reflektovaly poškozený stav. Zhoršení ekologického stavu nebylo zapříčiněno změnou vhodných habitatů nebo jejich úbytkem, ale především přítomností nepůvodních druhů, jež hodnotu indexu CZI výrazně snižují. Závěry našeho průzkumu poukazují na skutečnost, že k významným změnám ve společenstvu juvenilních ryb může docházet na téže lokalitě i ve velmi krátkém časovém úseku (jednoho roku). Meziroční změny mohou být proto značné, a z toho důvodu je důležité monitoring provádět každoročně.
ÚVOD
Velká část říčních systémů je silně pozměněna či poškozena lidskou činností [1], jako jsou nevhodné hydromorfologické úpravy i manipulace na vodních elektrárnách [2, 3], zavlečení invazních druhů, nadměrný vstup živin a znečištění cizorodými látkami [3]. Tyto multistresory významně ovlivňují celé vodní ekosystémy [1]. Voda a její kvalita hraje významnou roli z hlediska její využitelnosti jako nenahraditelné vstupní suroviny pro nespočet odvětví činností člověka [4]. Stejně tak i životní prostředí, na které je vázáno částečně nebo celým životním cyklem velké množství organismů. Využití vodních organismů (bioty) jako indikátoru ekologického stavu má opodstatněný význam [5]. Jejich fyziologická tolerance a ekologické preference úzce souvisejí s podmínkami prostředí, v němž žijí, a jsou schopny rychle reflektovat změny prostředí [6, 7]. Bioindikátory jsou široce využívány k poskytování užitečných informací o změnách či znečištění životního prostředí a odrážejí dlouhodobé vlivy/stresory, jež na organismy nepůsobí odděleně, nýbrž současně [8]. Většinou jsou metody hodnocení založeny na taxonomickém složení společenstva, které poskytuje informace o biologických interakcích, vnitřním utváření společenstva, ale i fungování daného ekosystému [9]. Společenstvo juvenilních ryb (tj. 0+, kde 0 znamená žádná prožitá zima a + znamená prožitá vegetační sezona) proto představuje vhodný nástroj pro monitoring ekologického stavu ve vodních tocích, zejména proto, že je většina českých a moravských vodních toků zarybňována, tj. jsou vysazováni subadultní a adultní jedinci [6]. Juvenilní ryby (0+) reflektují bezprostředně reprodukční úspěch nebo neúspěch v posledním období tření a vykazují výrazně rychlejší reakci na měnící se podmínky prostředí než dospělí jedinci [6, 10]. Kromě reprodukční úspěšnosti dospělců je společenstvo juvenilních ryb (0+) ovlivněno přežíváním jejich raných stadií, které jsou velmi těsně spjaty s výskytem vhodných mikro až mezohabitatů [11], jako jsou mělké úseky s dostatkem potravy a úkrytů, tzv. „rybí školky“ [10, 11]. Společenstvo juvenilních ryb (0+) je také utvářeno sezonními i mezisezonními změnami habitatů a hydrologickým [12] i teplotním režimem, který má podstatný vliv na celkovou diverzitu i abundanci jednotlivých druhů [13, 14]. Změny prostředí můžeme sledovat prostřednictvím diverzity v lokálním měřítku, a to na základě druhů v daném společenstvu (α diverzita) nebo i na širší škále, mezi jednotlivými společenstvy (β diverzita, [15, 16]). Cílem studie bylo posouzení společenstva juvenilních ryb (0+) a ekologického stavu vodních toků podle českého multimetrického indexu (CZI) v rámci jednotlivých povodí během let 2019–2021 na 22 lokalitách, které reprezentují závěrové profily a významné páteřní toky v ČR.
METODIKA
Biologické hodnocení sledovaných vodních toků bylo provedeno pomocí přirozeného rybího společenstva, tj. juvenilních ryb (0+). Metodika byla sestavena tak, aby bylo možné s jejím použitím provést odlov, základní zpracování a vyhodnocení vzorků ryb (0+) [17, 18]. Vybraná metodika reprezentuje současný stav vodních toků [19], kdy jsou odebírány pouze ryby, které jsou staré maximálně několik měsíců. Ichtyologický průzkum probíhal na 22 lokalitách (obr. 1), jež byly vybrány na základě předešlých poznatků z monitoringu jakosti vod, který provádí Český hydrometeorologický ústav [19]. Sledované lokality byly umístěny v závěrových profilech a na páteřních tocích ČR (obr. 1). Vzorkovací místa pro odlov juvenilních ryb (0+) byla umístěna pod obcemi a přilehlými aglomeracemi z důvodu možného ovlivnění technickými úpravami, jezovými manipulacemi, vypouštěním odpadních vod i plošnými zdroji znečištění, a to zejména ve významných zemědělských oblastech. Díky danému vzorkovacímu designu bylo možné objektivně zachytit vliv lidské činnosti na společenstvo juvenilních ryb (0+) mezi jednotlivými povodími, ale i napříč ČR.
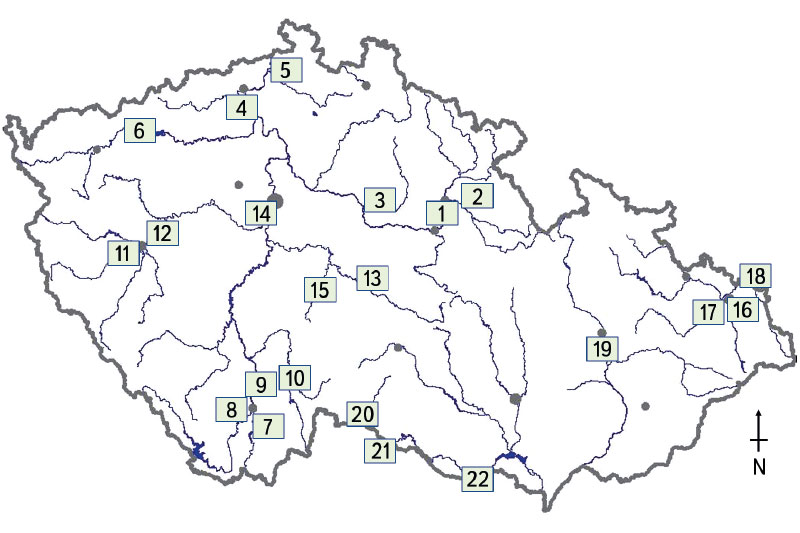
Obr. 1. Situační mapa sledovaných vodních toků s vyznačenými profily, na nichž probíhal odlov juvenilních ryb (0+): 1) Labe – Hradec Králové, 2) Orlice – Nepasice, 3) Cidlina – Sány, 4) Labe – Litoměřice, 5) Ploučnice – Děčín (Březiny), 6) Ohře – Želina, 7) Malše – Roudné, 8) Vltava – Boršov, 9) Vltava – Hluboká nad Vltavou, 10) Lužnice – Veselí nad Lužnicí, 11) Mže – Plzeň, 12) Berounka – Plzeň, 13) Sázava – Zruč nad Sázavou, 14) Vltava – Praha (Vrané), 15) Želivka – Poříčí, 16) Ostravice – Ostrava, 17) Odra – Ostrava (Svinov), 18) Olše – Věřňovice, 19) Morava – Blatec, 20) Moravská Dyje – Písečné, 21) Dyje – Podhradí, 22) Dyje – Jevišovka
Fig. 1. Map of rivers with sites where juvenile fish assemblages were monitored. 1) Labe – Hradec Králové, 2) Orlice – Nepasice, 3) Cidlina – Sány, 4) Labe – Litoměřice, 5) Ploučnice – Děčín (Březiny), 6) Ohře – Želina, 7) Malše – Roudné, 8) Vltava – Boršov, 9) Vltava – Hluboká nad Vltavou, 10) Lužnice – Veselí nad Lužnicí, 11) Mže – Plzeň, 12) Berounka – Plzeň, 13) Sázava – Zruč nad Sázavou, 14) Vltava – Praha (Vrané), 15) Želivka – Poříčí, 16) Ostravice – Ostrava, 17) Odra – Ostrava (Svinov), 18) Olše – Věřňovice, 19) Morava – Blatec, 20) Moravská Dyje – Písečné, 21) Dyje – Podhradí, 22) Dyje – Jevišovka
Odlov ryb
Odlovy ryb (0+) byly prováděny od druhé poloviny srpna do druhé poloviny září. Konec léta představuje vhodné období pro odběr vzorků juvenilních ryb (0+) z důvodu relativně nízkých a stabilních průtoků. I abundance juvenilních ryb (0+) je už relativně stabilní, ve srovnání s vysokou mortalitou, jež nastává během prvních týdnů až měsíců po vylíhnutí/vykulení [10]. Juvenilní ryby (0+) se v tomto období stále zdržují v mělkých úsecích podél břehu a ještě se nepřesouvají do hlubších částí vodních toků (na zimoviště), k čemuž dochází zpravidla během podzimních měsíců [10]. V tomto období jsou juvenilní ryby (0+) již dostatečně odrostlé, determinační znaky mají podobné dospělcům a jejich determinaci je možné provádět přímo v terénu [17, 20].
Odlov ryb byl prováděn podél břehové linie vodního toku (obr. 2) bateriovým elektrickým agregátem (typu SEN a LENA od firmy Bednář) s výstupní frekvencí 50–95 Hz [10, 21]. Ryby byly loveny pomocí stejnosměrného pulzního proudu, který není při daném rozmezí kmitočtu pro zdraví ryb nebezpečný [17, 20]. Délka proloveného úseku se odvíjela od množství mezohabitatů (mělké proudné úseky, mrtvé dřevo, vodní i zaplavená terestrická vegetace, stojatá voda) a pohybovala se v rozmezí od 50 m do 200 m (medián 100 m). Sledovaný úsek byl rozdělen na několik dílčích úseků tak, aby byla zachycena významná část variability prostředí a celkové společenstvo juvenilních ryb (0+). Následně po ulovení byly ryby determinovány přímo na dané lokalitě (obr. 3).

Obr. 2. Odlov juvenilních ryb v mělkých úsecích podél břehové linie
Fig 2. Juvenile fish assamblages sampling in shallow sections along the riverbank

Obr. 3. Determinace odlovených ryb v terénu
Fig. 3. The determination of juvenile fish
ZPRACOVÁNÍ DAT
Zhodnocení ekologického stavu sledovaných vodních toků bylo provedeno prostřednictvím českého multimetrického indexu (CZI), který kombinuje několik metrik, jejichž výsledky jsou spojeny do multimetrického výstupu a zahrnují několik atributů společenstva. Mezi metriky, jež popisují a hodnotí podmínky prostředí, patří nadmořská výška, řád vodního toku dle Strahlera, úmoří, typ toku (A – horské potoky až G – nížinné řeky) a typické taxony pro daný typ vodního toku i nepůvodní druhy, které významným způsobem snižují výslednou hodnotu indexu [16]. Multimetrický index byl vypočítán dle následující rovnice:

kde
wi je váha metriky při výpočtu CZI
TD počet typických taxonů
AR abundance reofilů (proudomilné druhy)
ND1 přítomnost nežádoucích druhů
ND2 relativní zastoupení nežádoucích druhů – nabývá hodnot od 0 do 1 (kategorie CZI, 0–0,2 zničený; > 0,2–0,4 poškozený; > 0,4–0,6 střední; > 0,6–0,8 dobrý a > 0,8–1 výborný). Horní a dolní mez hodnot metrik slouží k výpočtu Ecological Quality Ratio (EQR), tj. poměru mezi zjištěnými a očekávanými (referenčními) hodnotami [16].
Rozdíly ve společenstvu juvenilních ryb byly vyhodnoceny v programu R softwaru ver. 4.2.2 [21] prostřednictvím metody PERMANOVA (Permutational Multivariate Analysis of Variance) a zobrazeny pomocí vícenásobného škálování – NMDS (Non-Metric Multidimensional Scaling). Vizualizace rozdílů ve společenstvu ryb byla zobrazena prostřednictvím kódového označení jednotlivých druhů (AA – Alburnus alburnus, AB – Abramis brama, AN – Anguilla anguilla, AP – Alburnoides bipunctatus, AU – Leuciscus aspius, BB – Barbus barbus, BJ – Blicca bjoerkna, CA – Carassius gibelio, CG – Cottus gobio, CN – Chondrostoma nasus, CT – Cobitis taenia, CY – Cyprinus carpio, EL – Esox lucius, GA – Gasterosteus aculeatus, GC – Gymnocephalus cernua, GG – Gobio gobio, GL – Romanogobio albipinnatus, LC – Squalius cephalus, LG – Lepomis gibbosus, LI – Leuciscus idus, LL – Leuciscus Leuciscus, LT – Lota lota, NB – Barbatula barbatula, NM – Neogobius melanostomus, PF – Perca fluviatilis, PM – Proterorhinus semilunaris, PP – Phoxinus phoxinus, PR – Pseudorasbora parva, RR – Rutilus rutilus, RS – Rhodeus amarus, SE – Scardinius erythrophthalmus, SG – Silurus glanis, SL – Sander lucioperca, ST – Salmo trutta m. fario, TT – Tinca tinca, VV – Vimba vimba). Porovnání rozdílů ve společenstvu juvenilních ryb mezi jednotlivými lety (2019–2021) bylo provedeno s použitím euklidovské vzdálenosti (Jaccard index). Pro zhodnocení (beta) diverzity společenstva mezi jednotlivými lokalitami ve sledovaném období byl použit Cao index [22].
VÝSLEDKY
Společenstvo juvenilních ryb bylo poměrně bohaté, celkem bylo zaznamenáno 36 druhů na 22 lokalitách. Ve složení druhového společenstva mezi jednotlivými lokalitami byly značné rozdíly, minimálně byly zaznamenány čtyři druhy na lokalitu (Cidlina – Sány v roce 2019), naopak nejvíce druhů (15) bylo uloveno v roce 2021 na lokalitě Labe – Hradec Králové (úsek byl loven pod jezem v blízkosti obce Vysoká nad Labem). Ve sledovaném období bylo v průměru na lokalitách odloveno devět druhů (průměrně bylo zaznamenáno 7,1 druhu na lokalitu v roce 2019, 8,7 v roce 2020 a 9,7 v roce 2021). Mezi druhy s nejvyšší abundancí patřil jelec tloušť (Squalius cephalus ∑6156 ind. [individuum], obr. 4, tab. 1), hrouzek obecný (Gobio gobio ∑2976 ind., obr. 4, tab. 1), hořavka duhová (Rhodeus amarus ∑2518 ind., obr. 4, tab. 1), ouklej obecná (Alburnus alburnus ∑2447 ind., obr. 4, tab. 1), plotice obecná (Rutilus rutilus ∑2007 ind., obr. 4, tab. 1) a parma obecná (Barbus barbus ∑1434 ind., obr. 4, tab. 1). Jednotka lovného úsilí (CPUE [Catch Per Unit Effort]) mezi jednotlivými lety a lokalitami výrazně kolísala, minimální hodnota CPUE byla zaznamenána v roce 2019 na Berounce v Plzni (Bukovec) 0,3 ind.m-1 (obr. 5) a maximální 25,4 ind.m-1 na Olši ve Věřňovicích v roce 2021 (obr. 5).
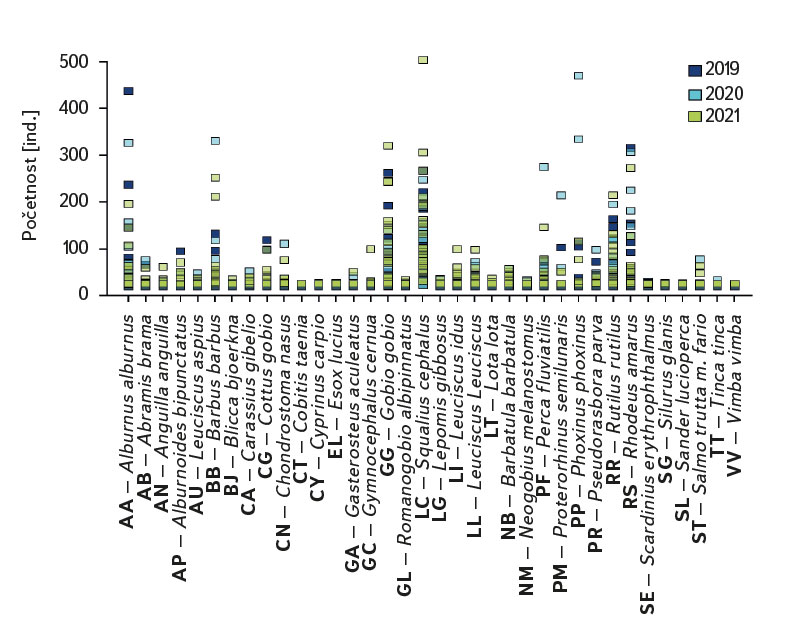
Obr. 4. Abundance jednotlivých druhů juvenilních ryb za období 2019–2021
Fig. 4. The results of juvenile fish survey, abundance of fish species between 2019–2021
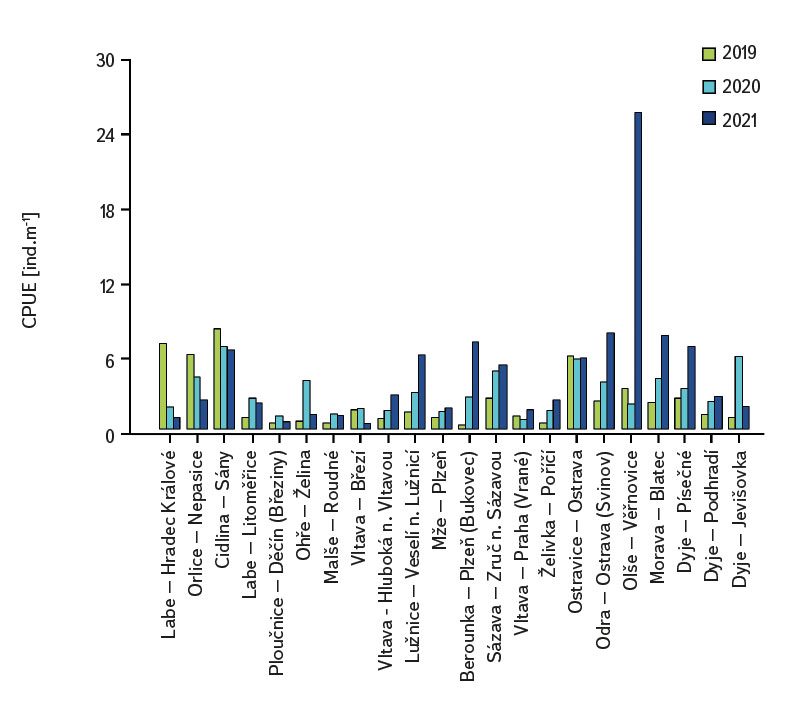
Obr. 5. Výsledky lovného úsilí (CPUE) na sledovaných lokalitách za období 2019–2021
Fig. 5. The results of catch per unit effort (CPUE) at monitored localities between 2019–2021
Tab. 1. Přehled ulovených juvenilních ryb (0+) na sledovaných lokalitách za období 2019–2021
Tab 1. Summary of juvenile fish (0+) caught at monitored sites for the period 2019–2021

Průměrná hodnota CPUE mezi lokalitami a lety dosahovala hodnot 3,3 ind.m-1. Průměrná a vyšší hodnota CPUE byla za sledované období zaznamenána ve 23 případech (v roce 2019 na pěti a shodně v letech 2020 a 2021 na devíti lokalitách). Podobně jako CPUE, tak i biomasa ryb vykazovala vysokou variabilitu mezi lety a lokalitami. Nejnižší hodnoty 0,4 g.m-1 byly zaznamenány v roce 2019 na Vltavě v Praze (Vrané, obr. 6), naopak nejvyšší 5,0 g.m-1 byly evidovány v roce 2021 na Odře v Ostravě (Svinov, obr. 6). Průměrná hodnota biomasy mezi lokalitami ve sledovaném období dosahovala 1,8 g.m-1. Průměrná a vyšší hodnota biomasy byla evidována ve 27 případech (v roce 2019 byla nalezena na sedmi, v roce 2020 na osmi a v roce 2021 na 12 lokalitách). Zhodnocení ekologického stavu podle českého multimetrického indexu (CZI) prokázalo podstatné změny na sledovaných lokalitách, které nastaly během let 2019–2021. Zhoršení stavu bylo zaznamenáno na čtyřech sledovaných lokalitách ve srovnání s předchozími lety (Labe – Hradec Králové, Ploučnice – Děčín/Březiny, Mže – Plzeň, Dyje – Podhradí, obr. 7). Nejnižší hodnoty CZI, a tedy i nejhorší ekologický stav (tj. zničený a poškozený) byly zaznamenány na lokalitách Ohře – Želina (0,200, obr. 7), Dyje – Jevišovka (0,295, obr. 7), Cidlina – Sány (0,305, obr. 7), a Dyje – Podhradí (0,344, obr. 7). Na Ohři v Želině v populaci dominoval zejména okoun říční (Perca fluviatilis), s minoritním podílem plotice obecné a koljušky tříostné (Gasterosteus aculeatus). Ve společenstvu na Dyji v Jevišovce majoritní podíl tvořily hlavačka poloměsíčitá (Proterorhinus semilunaris), hořavka duhová a plotice obecná. Na Cidlině v Sánech tvořili ve společenstvu majoritní podíl hořavka duhová, hrouzek obecný, plotice obecná a střevlička východní (Pseudorasbora parva). Na Dyji v Podhradí dominovali ve společenstvu ryb jelec tloušť, plotice obecná a okoun říční. Zlepšení ekologického stavu bylo detekováno celkem na devíti lokalitách (obr. 7). Nejvýraznější zlepšení za sledované období bylo zaznamenáno na čtyřech lokalitách, tj. na Labi v Litoměřicích, Lužnici ve Veselí n. Lužnicí, Vltavě v Praze (Vrané) a Dyji v Písečném. Na lokalitách Orlice v Nepasicích, Olše ve Věřňovicích ekologický stav nabýval 1. třídy (tj. velmi dobrý). U zbývajících lokalit byl stav spíše setrvalý, nedocházelo ani k výraznému zlepšení, ani ke zhoršení (obr. 7). Mnohorozměrné analýzy prokázaly významné rozdíly ve společenstvu juvenilních ryb v letech 2019–2021 (P = 0,011, obr. 8a), ale nebyly prokázány rozdíly v diverzitě mezi společenstvy napříč sledovanými lokalitami (P = 0,086, obr. 8b).
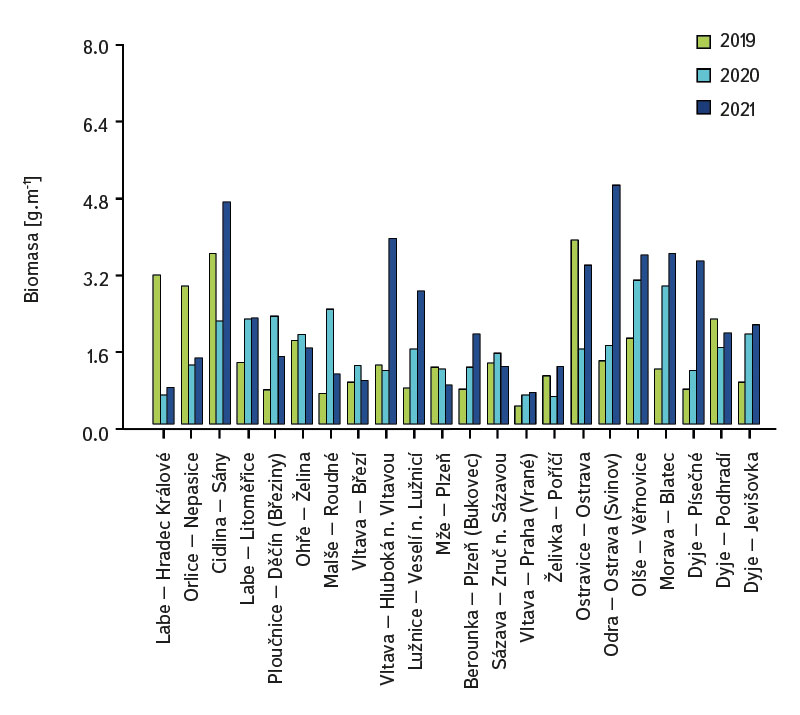
Obr. 6. Biomasa juvenilních ryb na sledovaných lokalitách za období 2019–2021
Fig. 6. Biomass of juvenile fish at monitored localities between 2019–2021
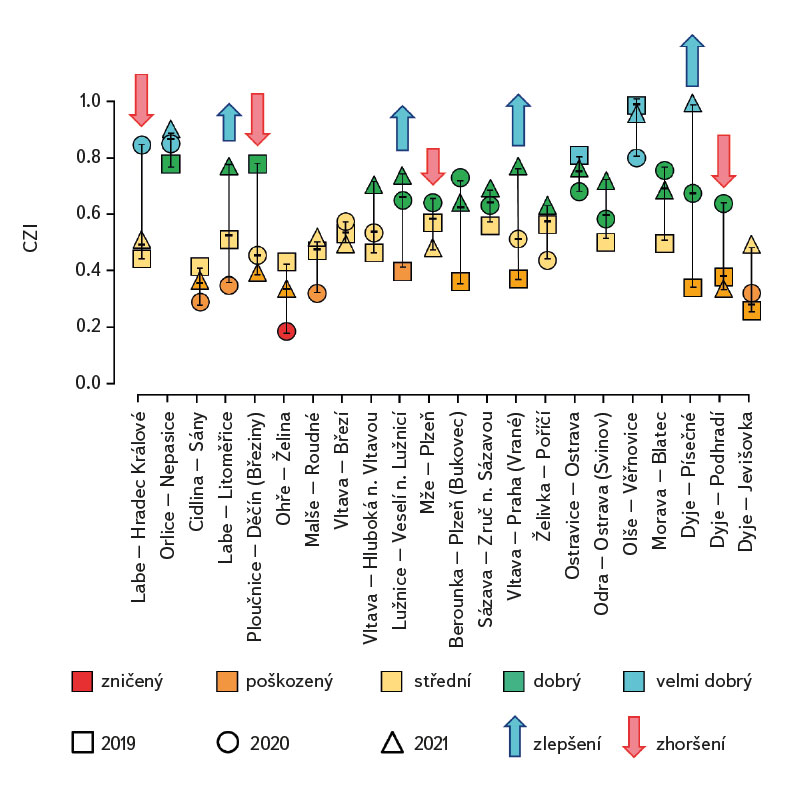
Obr. 7. Zhodnocení ekologického stavu na sledovaných lokalitách podle českého multimetrického indexu (CZI) za období 2019–2021
Fig. 7. The evaluation of ecological status using the Czech multimetric index (CZI) at monitored localities between 2019–2021
DISKUZE
Studie byla prováděna napříč ČR. Jednotlivé vodní toky a lokality se od sebe výrazně lišily nejen vodnatostí, geomorfologií, ale i technickými úpravami koryta. Na všech 22 sledovaných lokalitách bylo společenstvo juvenilních ryb velmi rozmanité. Druhová diverzita kolísala napříč jednotlivými lety i lokalitami, celkem bylo zaznamenáno 36 druhů (minimálně byly zachyceny čtyři a maximálně 15 druhů na lokalitu). Hodnoty CPUE vykazovaly poměrně vysokou variabilitu mezi lokalitami i sledovanými lety (obr. 5). Nejnižší hodnoty CPUE (0,3 ind.m-1, obr. 5) byly zaznamenány na Berounce v Plzni v roce 2019, ale v roce 2021 byly již evidovány hodnoty CPUE 6,8 ind.m-1 (obr. 5). Naopak nejvyšší hodnoty CPUE byly v roce 2021 na Olši ve Věřňovicích 25,4 ind.m-1, avšak v předchozích letech byly zaznamenány výrazně nižší abundance (4,5 a 2,1 ind.m-1, obr. 5). Obdobně i biomasa vykazovala velkou proměnlivost ve sledovaném období mezi lokalitami i lety, nejnižší hodnoty byly zjištěny na Vltavě v Praze (0,4 g.m-1, obr. 6) v roce 2019, ale v roce 2021 biomasa dosahovala téměř dvojnásobných hodnot (obr. 6). Naopak nejvyšší hodnoty 5,0 g.m-1 byly v roce 2021 na Odře v Ostravě (v městské části Svinov, obr. 6), nicméně v předchozích letech byly zaznamenány více než třikrát nižší hodnoty biomasy (obr. 6). Výrazné rozdíly v biomase i CPUE mezi jednotlivými lety v rámci téže lokality mohou souviset s meziročními rozdíly, výkyvy teplot či fluktuací hladiny (povodně, sucho), které mají výrazný vliv na reprodukční potenciál ryb i celé jejich společenstvo [6, 24, 25]. Rozdíly v abundanci i biomase mohou být taktéž ovlivněny meziročními biologickými cykly, jako jsou velikosti jednotlivých kohort, které vstupují do rozmnožování [6, 10], jež se mohou mezi jednotlivými lety výrazně lišit. Dále mohou být zapříčiněny i fluktuací dostupné potravy, tj. změnou ve společenstvu mikro a makrozoobentosu, které představuje významný zdroj potravy pro juvenilní ryby [24, 26, 27]. Mimo jiné i výrazná fluktuace teploty má citelný vliv na reprodukci ryb [13, 28], protože vyšší teploty vody mohou přispět k dřívější době tření, naopak náhlý pokles může zpomalit či oddálit tření ryb [13, 29].

Odlov juvenilních ryb v technicky silně upraveném korytě

Řeka Ohře u Želinského meandru

Výběr vhodného úseku toku v závislosti na variabilitě prostředí


Po determinaci byly ryby šetrně vypuštěny zpět do řeky
Lze se domnívat, že výrazný pokles teploty v jarním období může zapříčinit i absenci kohorty, zejména u ryb s dávkovým výtěrem, jako jsou např. jelec tloušť a ostroretka stěhovavá [30]. V roce 2020 byl únor a březen výrazně teplotně nadprůměrný, naopak květen byl teplotně velmi chladný (s odchylkou -2,1 °C od normálu, [31]). Na řadě lokalit byla evidována v tomto roce během odlovů ryb chybějící kohorta, resp. velikostní spektrum se pohybovalo pouze ve dvou kategoriích (cca 20–30 mm a 40–50 mm délky těla), střední kategorie 30–40 mm téměř chyběla (týkalo se to zejména jelce tlouště a ostroretky stěhovavé). I přes značné rozdíly v abundanci jednotlivých druhů (obr. 4) a výrazné variabilitě v druhové diverzitě (obr. 8a) i vzhledem k podstatným rozdílům mezi řadou lokalit (obr. 8b) nebyly nalezeny statisticky významné rozdíly ve společenstvu juvenilních ryb mezi sledovanými lokalitami (P = 0,086, obr. 8b), avšak tato hodnota je poměrně blízko k hladině významnosti (P = 0,05). Naopak byly zaznamenány významné rozdíly ve společenstvu v období 2019–2021 (P = 0,011, obr. 8a), kdy se druhová variabilita nápadně měnila během sledovaných let (obr. 8a). Neprůkazné rozdíly ve společenstvu juvenilních ryb mezi lokalitami mohly být zapříčiněny významným zastoupením eurytopních druhů, neboť sledované lokality se nalézají spíše v dolních částech vodních toků, a proto si společenstva mezi lokalitami mohla být i dosti podobná. Naopak významné rozdíly ve společenstvu ryb mezi sledovanými lety mohou poukazovat na zásadní změny, které se odehrávají během jednotlivých let, nebo mohly být zachyceny odrazy běžných meziročních fluktuací jinak stabilního společenstva [25]. Podle českého multimetrického indexu dvě lokality vykazovaly téměř stabilně nejlepší skladbu plůdkového společenstva, tj. výborný ekologický stav (0,863–1,0 CZI, obr. 7). Šlo o Orlici v Nepasicích a Olši ve Věřňovicích. Břehy a dno byly tvořeny středně hrubým štěrkem až pískem. Vyskytovalo se zde značné množství mezohabitatů, které byly vhodné jak pro reprodukci, tak i pro růst a přežívání plůdkového společenstva [32], tj. mělké pláže s malou rychlostí proudu a větším množstvím mrtvého dřeva, jež tvořily vhodný habitat s dostatkem potravy a úkrytů [10, 33]. Naopak nejnižší hodnoty CZI (0,200, 0,296, 0,305, 0,344, obr. 7), které představují „nejhorší“ (zničený až poškozený stav) byly nalezeny na Ohři v Želině, Dyji v Jevišovce, Cidlině v Sánech a Dyji v Podhradí. Ohře a Dyje byly ovlivněny přilehlými vodními nádržemi (Nechranická, Vranovská a Novomlýnská nádrž), do nichž tvoří hlavní přítoky. Současně nádrže ovlivňují i výsledné společenstvo juvenilních ryb (např. výškou vzdutí i rozmnožováním části obsádky nádrže v přítocích). Ve sledovaných úsecích bylo dno poměrně mělké, kamenité až písčité a jen místy porostlé řasovými nárosty a vodními makrofyty. Na Ohři bylo druhové společenstvo poměrně chudé, s převažující dominancí zejména okouna říčního, menším zastoupením plotice obecné a koljušky tříostné. V jarních měsících míří část obsádky z přehrady do přítoků, kde se rozmnožuje [34, 35]. V brzkých jarních měsících dochází k rozmnožování okouna [35] a následně plotice [36]. Okoun říční je schopen už ve velikosti od 25–30 mm aktivně lovit menší velikosti juvenilních ryb. Této velikosti běžně dorůstá v průběhu července až srpna [37–39]. Jeho velká abundance společně s enormním predačním potenciálem mu umožní převládnutí ve společenstvu juvenilních ryb, kde následně tvoří dominantní podíl. Nízké hodnoty českého multimetrického indexu na Dyji v Podhradí a v Jevišovce byly způsobeny relativně malou abundancí reofilních druhů, vyšší abundancí eurytopních druhů, jako byly plotice obecná a hořavka duhová, a zejména přítomností nepůvodních druhů, jako byli hlavačka poloměsíčitá, střevlička východní a karas stříbřitý (Carassius gibelio). Na Cidlině v Sánech ve společenstvu ryb v daném období klesala abundance hrouzka obecného a postupně začala dominovat plotice obecná a střevlička východní. Společenstvo bylo ovlivněno blízkostí Žehuňského rybníka, který měl vliv na průtokové poměry a teplotní režim a může sloužit i coby rezervoár nepůvodních druhů, jako je střevlička východní. Zhoršení ekologického stavu podle CZI během sledovaného tříletí bylo evidováno na čtyřech lokalitách (Labe – Hradec Králové, Ploučnice – Děčín/Březiny, Mže – Plzeň a Dyje – Podhradí, obr. 7). V daném období nebyla zaznamenána výrazná změna mezohabitatů na sledovaných lokalitách (např. technické úpravy koryta či nadměrné zarůstání koryta makrofyty z důvodu nízkých průtoků). Zhoršení bylo způsobeno zejména přítomností nepůvodních druhů, jež výrazným způsobem snižují hodnotu CZI. Tyto druhy expandují již z nově kolonizovaných území dále nebo jsou záměrně či nezáměrně rozšiřovány s násadou ryb [40, 41], popřípadě pocházejí z úniků z rybníků a jiných vodních ploch (produkční, okrasné rybníky a jezírka), které jsou situovány v horních částech povodí [42]. Naopak zlepšení stavu během let 2019–2021 bylo zaznamenáno na čtyřech lokalitách (Vltava – Hluboká nad Vltavou, Vltava – Vrané nad Vltavou, Želivka – Poříčí, Dyje – Písečné, obr. 7). Zlepšení může souviset s vytvořením vhodných mezohabitatů pro přežívání plůdku, které vznikly v důsledku výraznějších hydrologických událostí (tj. zvýšených vodních stavů), jež byly zaznamenány především v jarních i podzimních měsících roku 2020 (Český hydrometeorologický ústav, nepublikovaná data). Výrazné fluktuace vodních stavů mohou mít za následek hydromorfologické změny v korytě řek [43, 44], zejména pročištění koryt od jemného anorganického i organického materiálu (detritu), jež mohou přispět k vytvoření řady mezohabitatů [11]. Ty mohou být následně využitelné pro jednotlivá stadia juvenilních ryb (0+) [32, 45, 46].
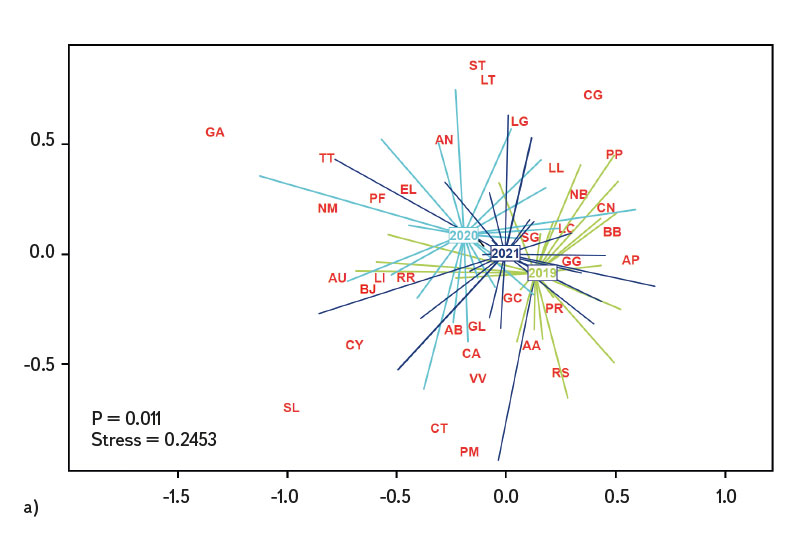
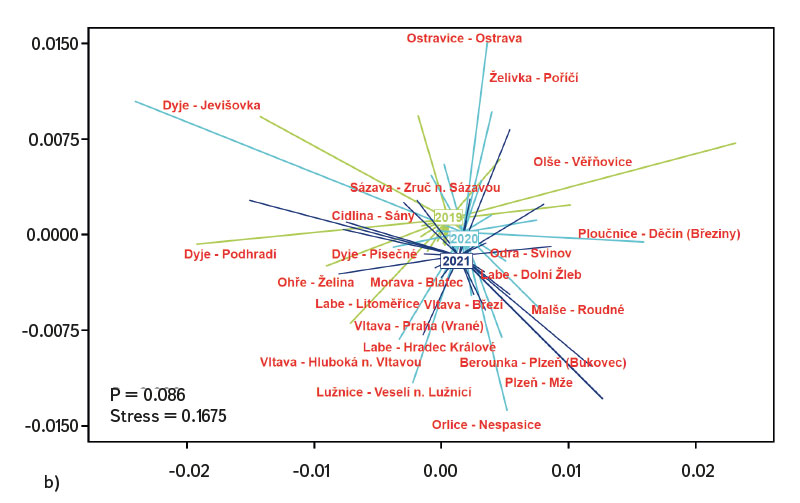
Obr. 8. Podobnosti mezi sledovanými lokalitami na základě společenstva juvenilních ryb, a) výsledky mnohorozměrné analýzy společenstva ryb mezi lety 2019–2021, b) rozdíly ve společenstvu ryb mezi sledovanými lokalitami v letech 2019–2021
Fig. 8. Similarities between monitored localities based on juvenile fish assemblages, a) the results of nonmetric multidimensional scaling of juvenile fish assemblages between 2019–2021, b) differences in juvenile fish assemblages between localities and across years 2019–2021
ZÁVĚR
Výsledky studie poukazují na skutečnost, že společenstvo juvenilních ryb (0+) představuje vhodný indikátor ekologického stavu našich vodních toků a je přímo i nepřímo ovlivněno přírodními podmínkami v daném roce. Zlepšení ekologického stavu bylo na mnoha lokalitách pravděpodobně způsobeno především zvýšenými vodními stavy, které působí jako významný korytotvorný prvek a jež zapříčinily odstranění nánosů a vytvoření vhodných mezohabitatů pro rozmnožování a následné přežívání prvních stadií juvenilních ryb (0+), zejména u reofilních druhů. Avšak zhoršení ekologického stavu nebylo zapříčiněno výraznou změnou vhodných habitatů nebo jejich náhlým úbytkem, nýbrž především přítomností nepůvodních druhů, jež hodnotu indexu CZI výrazně snižují. Závěry našeho průzkumu poukazují na skutečnost, že k významným změnám ve společenstvu juvenilních ryb může docházet na téže lokalitě i ve velmi krátkém časovém úseku (jednoho roku). Meziroční změny mohou být i velmi výrazné, a tak je důležité monitoring provádět každoročně, aby bylo možné „běžné“ fluktuace oddělit od zásadních změn probíhajících ve společenstvu juvenilních ryb (0+).
Příspěvek prošel recenzním řízením.
